Внешний вид рибосомы. Рибосомы эукариот и прокариот, сходства и различия
На заре развития жизни на Земле все клеточные формы были представлены бактериями. Они всасывали органические вещества, растворённые в первичном океане, через поверхность тела.
Со временем некоторые бактерии приспособились производить органические вещества из неорганических. Для этого они использовали энергию солнечного света. Возникла первая экологическая система, в которой эти организмы были производителями. В результате этого в атмосфере Земли появился кислород, выделяемый этими организмами. С его помощью можно из той же самой пищи получить гораздо больше энергии, а добавочную энергию использовать на усложнение строения тела: разделение тела на части.
Одно из важных достижений жизни — разделение ядра и цитоплазмы. В ядре находится наследственная информация. Специальная мембрана вокруг ядра позволила защитить от случайных повреждений. По мере необходимости цитоплазма получает из ядра команды, направляющие жизнедеятельность и развитие клетки.
Организмы, у которых ядро отделено от цитоплазмы, образовали надцарство ядерных (к ним относятся — растения, грибы, животные).
Таким образом, клетка — основа организации растений и животных — возникла и развилась в ходе биологической эволюции.
Даже не вооружённым глазом, а ещё лучше под лупой можно видеть, что мякоть зрелого арбуза состоит из очень мелких крупинок, или зёрнышек. Это клетки — мельчайшие «кирпичики», из которых состоят тела всех живых организмов, в том числе и растительных.
Жизнь растения осуществляется соединённой деятельностью его клеток, создающих единое целое. При многоклеточности частей растения существует физиологическое разграничение их функций, специализация различных клеток в зависимости от местоположения их в теле растения.
Растительная клетка отличается от животной тем, что имеет плотную оболочку, покрывающую внутреннее содержимое со всех сторон. Клетка не является плоской (как её принято изображать), она скорей всего похожа на очень маленький пузырёк, наполненный слизистым содержимым.
Строение и функции растительной клетки
Рассмотрим клетку как структурно-функциональную единицу организма. Снаружи клетка покрыта плотной клеточной стенкой, в которой имеются более тонкие участки — поры. Под ней находится очень тонкая плёнка — мембрана, покрывающая содержимое клетки — цитоплазму. В цитоплазме есть полости — вакуоли, заполненные клеточным соком. В центре клетки или около клеточной стенки расположено плотное тельце — ядро с ядрышком. От цитоплазмы ядро отделено ядерной оболочкой. По всей цитоплазме распределены мелкие тельца — пластиды.
Строение растительной клетки
Строение и функции органоидов растительной клетки
| Органоид | Рисунок | Описание | Функция | Особенности |
Клеточная стенка или плазматическая мембрана | Бесцветная, прозрачная и очень прочная | Пропускает в клетку и выпускает из клетки вещества. | Клеточная мембрана полупроницаемая |
|
Цитоплазма | Густое тягучее вещество | В ней располагаются все другие части клетки | Находится в постоянном движении |
|
Ядро (важная часть клетки) | Округлое или овальное | Обеспечивает передачу наследственных свойств дочерним клеткам при делении | Центральная часть клетки |
|
Сферической или неправильной формы | Принимает участие в синтезе белка | |||
 | Резервуар, отделённый от цитоплазмы мембраной. Содержит клеточный сок | Накапливаются запасные питательные вещества и продукты жизнедеятельности ненужные клетке. | По мере роста клетки мелкие вакуоли сливаются в одну большую (центральную) вакуоль |
|
Пластиды | Хлоропласты | Используют световую энергию солнца и создают органические из неорганических | Форма дисков, отграниченных от цитоплазмы двойной мембраной |
|
Хромопласты | Образуются в результате накопления каротиноидов | Жёлтые, оранжевые или бурые |
||
 | Лейкопласты | Бесцветные пластиды | ||
Ядерная оболочка | Состоит из двух мембран (наружная и внутренняя) с порами | Отграничивает ядро от цитоплазмы | Даёт возможность осуществляться обмену между ядром и цитоплазмой |
Живая часть клетки — это ограниченная мембраной, упорядоченная, структурированная система биополимеров и внутренних мембранных структур, участвующих в совокупности метаболических и энергетических процессов, осуществляющих поддержание и воспроизведение всей системы в целом.
Важной особенностью является то, что в клетке нет открытых мембран со свободными концами. Клеточные мембраны всегда ограничивают полости или участки, закрывая их со всех сторон.

Современная обобщенная схема растительной клетки
Плазмалемма (наружная клеточная мембрана) — ультрамикроскопическая плёнка толщиной 7,5 нм., состоящая из белков, фосфолипидов и воды. Это очень эластичная плёнка, хорошо смачивающаяся водой и быстро восстанавливающая целостность после повреждения. Имеет универсальное строение, т.е.типичное для всех биологических мембран. У растительных клеток снаружи от клеточной мембраны находится прочная, создающая внешнюю опору и поддерживающая форму клетки клеточная стенка. Она состоит из клетчатки (целлюлозы) — нерастворимого в воде полисахарида.
Плазмодесмы растительной клетки, представляют собой субмикроскопические канальцы, пронизывающие оболочки и выстланные плазматической мембраной, которая таким образом переходит из одной клетки в другую, не прерываясь. С их помощью происходит межклеточная циркуляция растворов, содержащих органические питательные вещества. По ним же идёт передача биопотенциалов и другой информации.
Порами называют отверстия во вторичной оболочке, где клетки разделяют лишь первичная оболочка и срединная пластинка. Участки первичной оболочки и срединную пластинку, разделяющие соседствующие поры смежных клеток, называют поровой мембраной или замыкающей пленкой поры. Замыкающую пленку поры пронизывают плазмодесменные канальцы, но сквозного отверстия в порах обычно не образуется. Поры облегчают транспорт воды и растворенных веществ от клетки к клетке. В стенках соседних клеток, как правило, одна против другой, образуются поры.
Клеточная оболочка имеет хорошо выраженную, относительно толстую оболочку полисахаридной природы. Оболочка растительной клетки продукт деятельности цитоплазмы. В её образовании активное участие принимает аппарат Гольджи и эндоплазматическая сеть.

Строение клеточной мембраны
Основу цитоплазмы составляет ее матрикс, или гиалоплазма, — сложная бесцветная, оптически прозрачная коллоидная система, способная к обратимым переходам из золя в гель. Важнейшая роль гиалоплазмы заключается в объединении всех клеточных структур в единую систему и обеспечении взаимодействия между ними в процессах клеточного метаболизма.
Гиалоплазма (или матрикс цитоплазмы) составляет внутреннюю среду клетки. Состоит из воды и различных биополимеров (белков, нуклеиновых кислот, полисахаридов, липидов), из которых основную часть составляют белки различной химической и функциональной специфичности. В гиалоплазме содержатся также аминокислоты, моносахара, нуклеотиды и другие низкомолекулярные вещества.
Биополимеры образуют с водой коллоидную среду, которая в зависимости от условий может быть плотной (в форме геля) или более жидкой (в форме золя), как во всей цитоплазме, так и в отдельных ее участках. В гиалоплазме локализуются и взаимодействуют между собой и средой гиалоплазмы различные органеллы и включения. При этом расположение их чаще всего специфично для определенных типов клеток. Через билипидную мембрану гиалоплазма взаимодействует с внеклеточной средой. Следовательно, гиалоплазма является динамической средой и играет важную роль в функционировании отдельных органелл и жизнедеятельности клеток в целом.
Цитоплазматические образования – органеллы
Органеллы (органоиды) — структурные компоненты цитоплазмы. Они имеют определённую форму и размеры, являются обязательными цитоплазматическими структурами клетки. При их отсутствии или повреждении клетка обычно теряет способность к дальнейшему существованию. Многие из органоидов способны к делению и самовоспроизведению. Размеры их настолько малы, что их можно видеть только в электронный микроскоп.
Ядро
Ядро — самая заметная и обычно самая крупная органелла клетки. Оно впервые было подробно исследовано Робертом Броуном в 1831 году. Ядро обеспечивает важнейшие метаболические и генетические функции клетки. По форме оно достаточно изменчиво: может быть шаровидным, овальным, лопастным, линзовидным.
Ядро играет значительную роль в жизни клетки. Клетка, из которой удалили ядро, не выделяет более оболочку, перестаёт расти и синтезировать вещества. В ней усиливаются продукты распада и разрушения, вследствие этого она быстро погибает. Образование нового ядра из цитоплазмы не происходит. Новые ядра образуются только делением или дроблением старого.
Внутреннее содержимое ядра составляет кариолимфа (ядерный сок), заполняющая пространство между структурами ядра. В нём находится одно или несколько ядрышек, а также значительное количество молекул ДНК, соединённых со специфическими белками — гистонами.

Строение ядра
Ядрышко
Ядрышко — как и цитоплазма, содержит преимущественно РНК и специфические белки. Важнейшая его функция заключается в том, что в нём происходит формирование рибосом, которые осуществляют синтез белков в клетке.
Аппарат Гольджи
Аппарат Гольджи — органоид, имеющий универсальное распространение во всех разновидностях эукариотических клеток. Представляет собой многоярусную систему плоских мембранных мешочков, которые по периферии утолщаются и образуют пузырчатые отростки. Он чаще всего расположен вблизи ядра.

Аппарат Гольджи
В состав аппарата Гольджи обязательно входит система мелких пузырьков (везикул), которые отшнуровываются от утолщённых цистерн (диски) и располагаются по периферии этой структуры. Эти пузырьки играют роль внутриклеточной транспортной системы специфических секторных гранул, могут служить источником клеточных лизосом.
Функции аппарата Гольджи состоят также в накоплении, сепарации и выделении за пределы клетки с помощью пузырьков продуктов внутриклеточного синтеза, продуктов распада, токсических веществ. Продукты синтетической деятельности клетки, а также различные вещества, поступающие в клетку из окружающей среды по каналам эндоплазматической сети, транспортируются к аппарату Гольджи, накапливаются в этом органоиде, а затем в виде капелек или зёрен поступают в цитоплазму и либо используются самой клеткой, либо выводятся наружу. В растительных клетках Аппарат Гольджи содержит ферменты синтеза полисахаридов и сам полисахаридный материал, который используется для построения клеточной оболочки. Предполагают, что он участвует в образовании вакуолей. Аппарат Гольджи был назван так в честь итальянского учёного Камилло Гольджи, впервые обнаружившего его в 1897 году.
Лизосомы
Лизосомы представляют собой мелкие пузырьки, ограниченные мембраной основная функция которых — осуществление внутриклеточного пищеварения. Использование лизосомного аппарата происходит при прорастании семени растения (гидролиз запасных питательных веществ).

Строение лизосомы
Микротрубочки
Микротрубочки — мембранные, надмолекулярные структуры, состоящие из белковых глобул, расположенных спиральными или прямолинейными рядами. Микротрубочки выполняют преимущественно механическую (двигательную) функцию, обеспечивая подвижность и сокращаемость органоидов клетки. Располагаясь в цитоплазме, они придают клетке определённую форму и обеспечивают стабильность пространственного расположения органоидов. Микротрубочки способствуют перемещению органоидов в места, которые определяются физиологическими потребностями клетки. Значительное количество этих структур расположено в плазмалемме, вблизи клеточной оболочки, где они участвуют в формировании и ориентации целлюлозных микрофибрилл оболочек растительных клеток.

Строение микротрубочки
Вакуоль
Вакуоль — важнейшая составная часть растительных клеток. Она представляет собой своеобразную полость (резервуар) в массе цитоплазмы, заполненную водным раствором минеральных солей, аминокислот, органических кислот, пигментов, углеводов и отделённую от цитоплазмы вакуолярной мембраной — тонопластом.
Цитоплазма заполняет всю внутреннюю полость только у самых молодых растительных клеток. С ростом клетки существенно изменяется пространственное расположение вначале сплошной массы цитоплазмы: у неё появляются заполненные клеточным соком небольшие вакуоли, и вся масса становится ноздреватой. При дальнейшем росте клетки отдельные вакуоли сливаются, оттесняя к периферии прослойки цитоплазмы, в результате чего в сформированной клетке находится обычно одна большая вакуоль, а цитоплазма со всеми органеллами располагаются около оболочки.
Водорастворимые органические и минеральные соединения вакуолей обусловливают соответствующие осмотические свойства живых клеток. Этот раствор определённой концентрации является своеобразным осмотическим насосом для регулируемого проникновения в клетку и выделения из неё воды, ионов и молекул метаболитов.
В комплексе со слоем цитоплазмы и её мембранами, характеризующимися свойствами полупроницаемости, вакуоль образует эффективную осмотическую систему. Осмотически обусловленными являются такие показатели живых растительных клеток, как осмотический потенциал, сосущая сила и тургорное давление.

Строение вакуоли
Пластиды
Пластиды — самые крупные (после ядра) цитоплазматические органоиды, присущие только клеткам растительных организмов. Они не найдены только у грибов. Пластиды играют важную роль в обмене веществ. Они отделены от цитоплазмы двойной мембранной оболочкой, а некоторые их типы имеют хорошо развитую и упорядоченную систему внутренних мембран. Все пластиды едины по происхождению.
Хлоропласты — наиболее распространённые и наиболее функционально важные пластиды фотоавтотрофных организмов, которые осуществляют фотосинтетические процессы, приводящие в конечном итоге к образованию органических веществ и выделению свободного кислорода. Хлоропласты высших растений имеют сложное внутреннее строение.
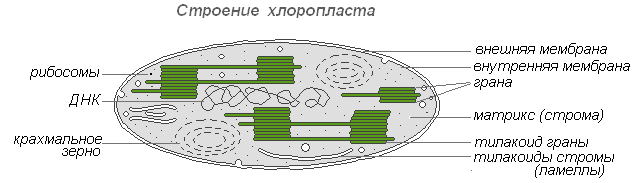
Строение хлоропласта
Размеры хлоропластов у разных растений неодинаковы, но в среднем диаметр их составляет 4-6 мкм. Хлоропласты способны передвигаться под влиянием движения цитоплазмы. Кроме того, под воздействием освещения наблюдается активное передвижение хлоропластов амебовидного типа к источнику света.
Хлорофилл — основное вещество хлоропластов. Благодаря хлорофиллу зелёные растения способны использовать световую энергию.
Лейкопласты (бесцветные пластиды) представляют собой чётко обозначенные тельца цитоплазмы. Размеры их несколько меньше, чем размеры хлоропластов. Более и однообразна и их форма, приближающая к сферической.

Строение лейкопласта
Встречаются в клетках эпидермиса, клубнях, корневищах. При освещении очень быстро превращаются в хлоропласты с соответствующим изменением внутренней структуры. Лейкопласты содержат ферменты, с помощью которых из излишков глюкозы, образованной в процессе фотосинтеза, в них синтезируется крахмал, основная масса которого откладывается в запасающих тканях или органах (клубнях, корневищах, семенах) в виде крахмальных зёрен. У некоторых растений в лейкопластах откладываются жиры. Резервная функция лейкопластов изредка проявляется в образовании запасных белков в форме кристаллов или аморфных включений.
Хромопласты в большинстве случаев являются производными хлоропластов, изредка — лейкопластов.

Строение хромопласта
Созревание плодов шиповника, перца, помидоров сопровождается превращением хлоро- или лейкопластов клеток мякоти в каратиноидопласты. Последние содержат преимущественно жёлтые пластидные пигменты — каратиноиды, которые при созревании интенсивно синтезируются в них, образуя окрашенные липидные капли, твёрдые глобулы или кристаллы. Хлорофилл при этом разрушается.
Митохондрии
Митохондрии — органеллы, характерные для большинства клеток растений. Имеют изменчивую форму палочек, зёрнышек, нитей. Открыты в 1894 году Р. Альтманом с помощью светового микроскопа, а внутреннее строение было изучено позднее с помощью электронного.

Строение митохондрии
Митохондрии имеют двухмембранное строение. Внешняя мембрана гладкая, внутренняя образует различной формы выросты — трубочки в растительных клетках. Пространство внутри митохондрии заполнено полужидким содержимым (матриксом), куда входят ферменты, белки, липиды, соли кальция и магния, витамины, а также РНК, ДНК и рибосомы. Ферментативный комплекс митохондрий ускоряет работу сложного и взаимосвязанного механизма биохимических реакций, в результате которых образуется АТФ. В этих органеллах осуществляется обеспечение клеток энергией — преобразование энергии химических связей питательных веществ в макроэргиеские связи АТФ в процессе клеточного дыхания. Именно в митохондриях происходит ферментативное расщепление углеводов, жирных кислот, аминокислот с освобождением энергии и последующим превращением её в энергию АТФ. Накопленная энергия расходуется на ростовые процессы, на новые синтезы и т. д. Митохондрии размножаются делением и живут около 10 дней, после чего подвергаются разрушению.
Эндоплазматическая сеть
Эндоплазматическая сеть — сеть каналов, трубочек, пузырьков, цистерн, расположенных внутри цитоплазмы. Открыта в 1945 году английским учёным К. Портером, представляет собой систему мембран, имеющих ультрамикроскопическое строение.

Строение эндоплазматической сети
Вся сеть объединена в единое целое с наружной клеточной мембраной ядерной оболочки. Различают ЭПС гладкую и шероховатую, несущую на себе рибосомы. На мембранах гладкой ЭПС находятся ферментные системы, участвующие в жировом и углеводном обмене. Этот тип мембран преобладает в клетках семян, богатых запасными веществами (белками, углеводами, маслами), рибосомы прикрепляются к мембране гранулярной ЭПС, и во время синтеза белковой молекулы полипептидная цепочка с рибосомами погружается в канал ЭПС. Функции эндоплазматической сети очень разнообразны: транспорт веществ как внутри клетки, так и между соседними клетками; разделение клетки на отдельные секции, в которых одновременно проходят различные физиологические процессы и химические реакции.
Рибосомы
Рибосомы — немембранные клеточные органоиды. Каждая рибосома состоит из двух не одинаковых по размеру частичек и может делиться на два фрагмента, которые продолжают сохранять способность синтезировать белок после объединения в целую рибосому.

Строение рибосомы
Рибосомы синтезируются в ядре, затем покидают его, переходя в цитоплазму, где прикрепляются к наружной поверхности мембран эндоплазматической сети или располагаются свободно. В зависимости от типа синтезируемого белка рибосомы могут функционировать по одиночке или объединяться в комплексы — полирибосомы.
Рибосомы - немембранные универсальные органеллы, в состав которых входят рРНК и белки. Открытые в 1955 году Джорджем Палладе. О важности этих органелл в клетке свидетельствует тот факт, что в 2009 году американские ученые В. Рамакришнан, Т. Стейц и А. Йонат за изучение структуры рибосом получили Нобелевскую премию по химии.
В клетке созревшие рибосомы находятся преимущественно в компартментах, где активно осуществляется биосинтез белков. Они могут быть свободно расположенными в цитоплазме, прикрепленными к мембранам зернистой ЭПС, на ядерной оболочке, в пластидах и митохондриях. Находятся в прокариотических и эукариотических клетках, за исключением эритроцитов млекопитающих. Учитывая массу и распространения различают два вида рибосом:
1) малые рибосомы (70S) - содержатся в клетках прокариот, а также в пластидах и митохондриях эукариот; такие рибосомы подключен к мембранам и имеют диаметр 15 нм;
2) большие рибосомы (80S) - содержатся в цитоплазме клеток эукариотического типа; такие рибосомы имеют диаметр около 22 нм и связанные с мембранами гранулярной ЭПС.
Строение . Структурная организация рибосом принципиально одинакова. Каждая из этих органелл состоит из двух субъединиц: большой и малой. Субъединицы рибосом, как правило, обозначаются единицами Сведберга (S), является мерой скорости седиментации при центрифугирования, и зависят от массы, размера и формы частицы. В рибосомах эукариот эти большая и малая субъединицы имеют константу седиментации Сведберга, соответственно, 60S и 40S. Сочетаются обе субъединицы поперечными сторонами с помощью ионов магния (Мд2 +) с образованием узкой щели. Рибосомы в эукариот синтезируются в ядрышке. Матрицей для рРНК есть участки ДНК. В прокариот рибосомы образуются в цитоплазме в результате простого сочетания компонентов.
Химическая организация. Рибосомы содержат рибосомальной РНК (рРНК) и белок: 40 60% рРНК и 60-40% белка. В рибосомах находится около 80-90% всей РНК клетки. Каждая субъединица содержит по одной или две молекулы рРНК в виде клубка, плотно упакованного белками, створююе рибонуклеопротеидний комплекс. При снижении концентрации ионов магния в растворе может наступить изменение конформации РНК и развертывания тяжа. Неработающие рибосомы постоянно обмениваются субъединицами. Собираются они только в момент синтеза белков и формируют вместе с иРНК полисомы, или полирибосомы. Рибосомы могут размещаться в цитоплазме клетки одиночно, тогда они функционально неактивны. Сбор рибосом на иРНК происходит в начале синтеза белка. Количество рибосом зависит от метаболической активности клетки. Особенно много полисом есть в клетках, которые быстро делятся, и в тех, которые продуцируют большое количество белков. Количество рибосом в таких клетках может достичь 50 000, что составляет около 25% массы всей клетки.
Функции . Методом меченых аминокислот обнаружено, что в рибосомах происходит синтез белков. Полипептидные молекулы белка синтезируются таким образом, что определенные аминокислоты в рибосоме соединяются друг с другом в соответствующей последовательности. Поэтому информационная РНК, кодирующей порядок размещения аминокислот, имеет перемещаться по рибосоме. Чем больше рибосом содержит полисома, тем больше молекул полипептидов будет синтезироваться на ней одновременно. Синтез белка на рибосомах начинается с прикрепления рибосомы к определенному участку иРНК.
Органеллы клетки, состоящие из белков и РНК и отвечающие за синтез белков, называются рибосомами. Количество рибосом в одной клетке сильно варьирует в зависимости от потребностей и может достигать нескольких миллионов.
Строение
Важнейшей органеллой клетки является ядро. Оно содержит генетическую информацию и ядрышко, где образуются рибосомы. Синтезированные рибосомы через поры ядерной мембраны попадают либо на эндоплазматическую сеть, либо в цитоплазму. В зависимости от расположения в эукариотической клетке выделяют два вида рибосом:
- связанные - располагаются на эндоплазматической сети (шероховатый вид);
- свободные - располагаются в цитозоле.
Гладкая ЭПС образуется после освобождения от рибосом. В растительных клетках гладкая ЭПС формирует провакуоли, из которых затем образуются вакуоли.

Рис. 1. Расположение рибосом в клетке.
Рибосомы - немембранные органеллы, имеющие округлую форму и состоящие из двух частей - субъединиц (большой и малой), каждая из которых представляет собой смесь рибосомальной РНК (рРНК) и белков. С химической точки зрения рибосома - нуклеопротеид, состоящий из нуклеиновых кислот и протеинов.

Рис. 2. Строение рибосом.
Связанные и свободные рибосомы называются цитоплазматическими рибосомами. Также существуют собственные рибосомы митохондрий и пластид. Они отличаются меньшим количеством белков и рРНК.
Различают четыре разновидности молекул РНК рибосомы:
ТОП-3 статьи которые читают вместе с этой
- 18S-РНК - содержит 1900 нуклеотидов;
- 5S-РНК - содержит 120 нуклеотидов;
- 5,8S-РНК - состоит из 160 нуклеотидов;
- 28S-РНК - состоит из 4800 нуклеотидов.
Малая частица рибосомы образована 30-35 белками и 18S-РНК. В большую субчастицу входит 45-50 белков и 5S-, 5,8S-, 28S-РНК.
В нерабочем состоянии части рибосом разъединены. Они соединяются с помощью информационной (матричной) РНК, обхватывая её с двух сторон. При синтезе белка рибосомы объединяются, образуя комплексы - полисомы или полирибосомы, связанные мРНК и напоминающие бусины на нитке.
Рибосомы прокариот меньше, чем эукариот. Диаметр рибосом клетки человека, животных, растений и грибов - 25-30 нм, бактерий - 15-20 нм.
Синтез белка
Главная функция рРНК - синтез белка и аминокислот.
Биосинтез белков включает два процесса:
- транскрипцию;
- трансляцию.
Транскрипция происходит с участием ДНК. Генетическую информацию считывает фермент РНК-полимераза, образуя мРНК. Далее начинается процесс трансляции, происходящий на рибосомах.
Этот процесс разделяется на три этапа:
- инициацию - начало синтеза;
- элонгацию - биосинтез;
- терминацию - завершение синтеза, отделение рибосомы.
При инициации происходит сборка рибосомы. Контактные части субъединиц называются активными центрами, между которыми располагается :
- мРНК в качестве «шаблона» синтеза;
- тРНК, осуществляющая перенос аминокислот на синтезируемую цепь;
- синтезируемый пептид, состоящий из аминокислот.
В процессе элонгации происходит удлинение полипептидной цепи за счёт присоединения аминокислот. Цепь отсоединяется от рибосомы на стадии терминации благодаря стоп-кодону - единицы генетического кода, шифрующего прекращение синтеза белка.

Рис. 3. Общая схема синтеза белка на рибосоме.
Биосинтез требует энергетических затрат. При присоединении одной аминокислоты расходуется по две молекулы АТФ (аденозинтрифосфата) и ГТФ (гуанозинтрифосфата). Кроме того, ГТФ тратится на процессы инициации и терминации.
Что мы узнали?
Из урока 9 класса кратко узнали о строении и функции рибосомы. Это важные органоиды клетки, осуществляющие биосинтез белка путём считывания информации с мРНК. Рибосомы образованы двумя частями (большой и малой), каждая из которых состоит из рибонуклеиновой кислоты и белков.
Тест по теме
Оценка доклада
Средняя оценка: 4.6 . Всего получено оценок: 110.
Рибосома(от «РНК» и soma – тело) – клеточный немембранный органоид, осуществляющий трансляцию (считывание кода мРНК и синтез полипептидов).
Рибосомы эукариот расположены на мембранах эндоплазматической сети (гранулярная ЭС) и в цитоплазме. Прикрепленные к мембранам рибосомы синтезируют белок «на экспорт», а свободные рибосомы – для нужд самой клетки. Различают 2 основных типа рибосом – прокариотные и эукариотные. В митохондриях и хлоропластах также имеются рибосомы, которые близки к рибосомам прокариот.
Рибосома состоит из двух субъединиц – большой и малой. У прокариотических клеток они обозначены 50S и 30S субъединицы, у эукариотических – 60S и 40S. (S – коэффициент, который характеризует скорость осаждения субъединицы при ультрацентрифугировании). Субъединицы эукариотических рибосом образуются путем самосборки в ядрышке и через поры ядра поступают в цитоплазму.
Рибосомы в клетках эукариот состоят из четырех нитей РНК (три молекулы рРНК в большой субъединице и одна молекула рРНК – в малой) и примерно 80 разных белков, т.е представляют собой сложнейший комплекс из молекул, скрепленных слабыми, нековалентными связями. (Рибосомы в клетках прокариот состоят из трех нитей РНК; две нити рРНК находятся в большой субъединице и одна рРНК – в малой). Процесс трансляции (биосинтеза белка) начинается со сборки активной рибосомы. Этот процесс называется инициацией трансляции. Сборка происходит строго упорядоченным образом, что обеспечивается функциональными центрами рибосом. Все центры находятся на контактирующих поверхностях обеих субъединиц рибосомы. Каждая рибосома работает как большая биохимическая машина, а точнее, как суперфермент, который, во-первых, правильно ориентирует участников (мРНК и тРНК) процесса друг относительно друга, а во-вторых, катализирует реакции между аминокислотами.
Активные центры рибосом:
1)центр связывания мРНК (М-центр);
2) пептидильный центр (П-центр). С этим центром в начале процесса трансляции связывается инициирующая тРНК; на последующих стадиях трансляции из А-центра в П-центр перемещается тРНК, удерживающая синтезированную часть пептидной цепи;
3)аминокислотный центр (А-центр) – место связывания кодона мРНК с антикодоном тРНК, несущей очередную аминокислоту.
4)пептидилтрансферазный центр (ПТФ-центр): он катализирует реакцию связывания аминокислот. При этом образуется еще одна пептидная связь, и растущий пептид удлиняется на одну аминокислоту.

Схема синтеза белка на рибосомах гранулярной эндоплазматической сети.
(рис. из книги биология клетки, том II )

Схематическое изображение полирибосомы. Синтез белка начинается со связывания малой субчастицы, в месте расположения AUG -кодона в молекуле информационной (матричной РНК) (рис. из книги биология клетки, том II ).
Эндоплазматическая сеть
Эндоплазматическая сеть (син. эндоплазматический ретикулум) – органоид эукариотической клетки. В клетках разного типа и при различных функциональных состояниях этот компонент клетки может выглядеть по-разному, но во всех случаях – это лабиринтная протяженная замкнутая мембранная структура, построенная из сообщающихся трубкообразных полостей и мешочков, называемых цистернами. Снаружи от мембран эндоплазматической сети находится цитозоль (гиалоплазма, основное вещество цитоплазмы), а просвет эндоплазматической сети представляет собой замкнутое пространство (компартмент), сообщающееся посредством везикул (транспортных пузырьков) с комплексом Гольджи и внешней для клетки средой. Эндоплазматическая сеть делится на две функционально различные структуры: гранулярную (шероховатую) эндоплазматическую сеть и гладкую(агранулярную) эндоплазматическую сеть.
Гранулярная эндоплазматическая сеть, в клетках секретирующих белок, представлена системой многочисленных плоских мембранных цистерн с рибосомами на наружной поверхности. Комплекс мембран гранулярной эндоплазматической сети связан с наружной мембраной оболочки ядра и перинуклеарной (околоядерной) цистерной.
В гранулярной эндоплазматической сети происходит синтез белков и липидов для всех мембран клетки, синтезируются ферменты лизосом, а также осуществляется синтез секретируемых белков, т.е. предназначенных для экзоцитоза. (Остальные белки синтезируются в цитоплазме на рибосомах, не связанных с мембранами ЭС.) В просвете гранулярной ЭС белок окружается мембраной, и образующиеся пузырьки отделяются (отпочковываются) от несодержащих рибосомы областей ЭС, которые и доставляют содержимое в другую органеллу – комплекс Гольджи – путем слияния с ее мембраной.
Та часть ЭС, на мембранах которой рибосомы отсутствуют, называется гладким эндоплазматическим ретикулумом. Гладкая эндоплазматическая сеть не содержит уплощенных цистерн, а представляет собой систему анастомозирующих мембранных канал
ов, пузырьков и трубочек. Гладкая сеть является продолжением гранулярной, однако не содержит рибофоринов – гликопротеиновых рецепторов, с которыми соединяется большая субъединица рибосом и поэтому не связана с рибосомами.
Функции гладкой эндоплазматической сети многообразны и зависят от типа клеток. Гладкая эндоплазматическая сеть участвует в метаболизме стероидных, например, половых гормонов. В ее мембранах локализованы управляемые кальциевые каналы и энергозависимые кальциевые насосы. Цистерны гладкой эндоплазматической сети специализированы для накопления в них Са 2+ путем постоянного откачивания Са 2+ из цитозоля. Подобные депо Са 2+ существуют в скелетной и сердечной мышцах, нейронах, яйцеклетке, эндокринных клетках и др. Различные сигналы (например, гормоны, нейромедиаторы, факторы роста) влияют на активность клеток путем изменения концентрации внутриклеточного посредника – Са 2+ . В гладкой эндоплазматической сети клеток печени происходит обезвреживание вредных веществ, (например ацетальдегида, образующегося из алкоголя), метаболическая трансформация лекарств, образование большей части липидов клетки и их накопление, например при жировой дистрофии. В полости ЭС содержится много различных молекул-компонентов. Среди них имеют большое значение белки шапероны.

Шапероны (англ. букв. – пожилая дама, сопровождающая молодую девушку на балах) – семейство специализированных внутриклеточных белков, обеспечивающих быстрое и правильное сворачивание (фолдинг) вновь синтезированных молекул белка. Связывание с шаперонами препятствует агрегации с другими белками и тем самым создает условия для формирования вторичной и третичной структуры растущего пептида. Шапероны принадлежат к трем белковым семействам, так называемым белкам теплового шока (hsp 60, hsp 70, hsp 90). Синтез этих белков активируется при многих стрессах, в частности, при тепловом шоке (отсюда и название h eart shook protein – белок теплового шока, а цифра обозначает его молекулярную массу в килодальтонах). Эти шапероны предотвращают денатурацию белков при высокой температуре и др. экстремальных факторах. Связываясь с аномальными белками, восстанавливают их нормальную конформацию и тем самым повышают выживаемость организма при резком ухудшении физико-химических параметров среды.
Рибосома представляет собой элементарную клеточную машину синтеза любых белков клетки. Все они построены в клетке одинаково, имеют одинаковую молекулярную композицию, выполняют одинаковую функцию – синтез белка – поэтому их можно так же считать клеточными органоидами. В отличие от других органоидов цитоплазмы (пластид, митохондрий, клеточного центра, мембранной вакуолярной системы и др.) они представлены в клетке огромным числом: за клеточный цикл их образуется 1 х 10 7 штук. Поэтому основная масса клеточной РНК представляет собой именно рибосомную РНК. РНК рибосом относительно стабильна, рибосомы могут существовать в клетках культуры ткани в течение нескольких клеточных циклов. В печеночных клетках время полужизни рибосом составляет 50-120 часов.
Рибосомы – это сложные рибонуклеопротеидные частицы, в состав которых входит множество молекул индивидуальных (неповторенных) белков и несколько молекул РНК, Рибосомы прокариот и эукариот по своим размерам и молекулярным характеристикам отличаются, хотя и обладают общими принципами организации и функционирования. К настоящему времени методом рентгеноструктурного анализа высокого разрешения полностью расшифрована структура рибосом.
Полная, работающая рибосома, состоит из двух неравных субъединиц, которые легко обратимо диссоциируют на большую субъединицу и малую. Размер полной прокариотической рибосомы составляет 20 х 17 х 17 нм, эукариотической – 25 х 20 х 20. Полная прокариотическая рибосома имеет коэффициент седиментации 70S и диссоциирует на две субъединицы: 50S и 30S. Полная эукариотическая рибосома, 80S рибосома, диссоциирует на 60S и 40S субъединицы. Форма и детальные очертания рибосом из разнообразных организмов и клеток, включая как прокариотические, так и эукариотические, поразительно похожи, хотя и отличаются рядом деталей. Малая рибосомная субъединица имеет палочковидную форму с несколькими небольшими выступами (см. рис. 81), ее длина составляет около 23 нм, а ширина – 12 нм. Большая субъединица похожа на полусферу с тремя торчащими выступами. При ассоциации в полную 70S рибосому малая субчастица ложится одним концом на один из выступов 50S частицы, а другим в ее желобок. В состав малых субъединиц входит по одной молекуле РНК, а в состав большой – несколько: у прокариот – две, а у эукариот – 3 молекулы. Характеристики молекулярной композиции рибосом даны в таблице 9.
Таблица 9. Молекулярная характеристика рибосом
Таким образом в состав эукариотической рибосомы входят четыре молекулы РНК разной длины: 28S РНК содержит 5000 нуклеотидов, 18SРНК – 2000, 5,8S РНК – 160, 5SРНК – 120.Рибосомные РНК обладают сложной вторичной и третичной структурой, образуя сложные петли и шпильки на комплементарных участках, что приводит к самоупаковке, самоорганизации этих молекул в сложное по форме тело. Так, например, сама по себе молекула 18S РНК в физиологических ионных условиях образует палочковидную частицу, определяющую форму малой субъединицы рибосом.
Под действием низких ионных сил, особенно при удалении ионов магния, плотные рибосомные субъединицы могут разворачиваться в рыхлые рибонуклеопротеидные тяжи, где можно наблюдать кластеры отдельных белков, но правильных структур, типа нуклеосом, нет, т.к. нет групп из сходных белков: в рибосоме все 80 белков разные.
Для того, чтобы образовались рибосомы необходимо наличие четырех типов рибосомных РНК в эквимолярных отношениях и наличие всех рибосомных белков. Сборка рибосом может происходить спонтанно in vitro , если последовательно добавлять к РНК белки в определенной последовательности.
Следовательно для биосинтеза рибосом необходим синтез множества специальных рибосомных белков и 4-х типов рибосомной РНК. Где эта РНК синтезируется, на каком количестве генов, где эти гены локализованы, как они организованы в составе ДНК хромосом – все эти вопросы в последние десятилетия были успешно разрешены при изучении строения и функции ядрышек.


