Регуляция сна и бодрствования. Современные представления о регуляции сна и бодрствования Суточные ритмы установлены в организме раз и навсегда

 Десинхронизация ритмов организма у человека, живущего в пещере глубоко под землей в изоляции от внешнего мира и использующего только искусственное освещение. Темные полосы - периоды сна, светлые - бодрствование.
Десинхронизация ритмов организма у человека, живущего в пещере глубоко под землей в изоляции от внешнего мира и использующего только искусственное освещение. Темные полосы - периоды сна, светлые - бодрствование.
 Супрахиазматическое ядро расположено над зрительным перекрестом в основании гипоталамуса. Оно получает вход от зрительной системы и отвечает за восприятие светлого времени суток как времени бодрствования, а также поддерживает суточный ритм.
Супрахиазматическое ядро расположено над зрительным перекрестом в основании гипоталамуса. Оно получает вход от зрительной системы и отвечает за восприятие светлого времени суток как времени бодрствования, а также поддерживает суточный ритм.
 Бодрствование - состояние психики, характеризующееся достаточно высоким уровнем электрической активности мозга, свойственным активному взаимодействию индивида с внешним миром. В поддержании состояния бодрствования важнейшую роль играет ретикулярная формация среднего мозга, от нейронов которой восходящие влияния идут к неспецифическим ядрам таламуса, а от них ко всем зонам коры больших полушарий. Бодрствование (обычное состояние не спящего человека) образует поле всевозможных сочетаний функций сознания - от состояния спокойного бодрствования через активное, напряженное бодрствование до выраженных аффектов.
Бодрствование - состояние психики, характеризующееся достаточно высоким уровнем электрической активности мозга, свойственным активному взаимодействию индивида с внешним миром. В поддержании состояния бодрствования важнейшую роль играет ретикулярная формация среднего мозга, от нейронов которой восходящие влияния идут к неспецифическим ядрам таламуса, а от них ко всем зонам коры больших полушарий. Бодрствование (обычное состояние не спящего человека) образует поле всевозможных сочетаний функций сознания - от состояния спокойного бодрствования через активное, напряженное бодрствование до выраженных аффектов.


 Сон – это физиологическое состояние, при котором значительно снижается реакция нервной системы и всего организма на внешние раздражители. Состояние сна является следствием включения особых центров сна головного мозга и сопровождается снижением интенсивности физиологических процессов, общей неподвижностью.
Сон – это физиологическое состояние, при котором значительно снижается реакция нервной системы и всего организма на внешние раздражители. Состояние сна является следствием включения особых центров сна головного мозга и сопровождается снижением интенсивности физиологических процессов, общей неподвижностью.
 Выделяют две основные стадии сна - «медленный» и «быстрый» сон. Феномен «быстрого» сна открыли в 1953 г. американские ученые А. Азеринский и К. Клейтман. Медленноволновой сон человека периодически прерывается короткими периодами низкоамплитудной ЭЭГ (похожей на десинхронизацию ЭЭГ при пробуждении), сопровождающимися быстрыми движениями глазных яблок. Отсюда часто употребляемое название этой стадии - парадоксальная, или стадия быстрых движений глаз (БДГ-сон, или REM-стадия - от rapid eye movement). Люди, разбуженные во время парадоксальной стадии сна, в 80 % случаев сообщали о сновидениях. Мышцы тела расслаблены сильнее всего во время парадоксального сна, однако вегетативные показатели (дыхание, сердцебиение, кровяное давление и др.) во время парадоксального сна могут соответствовать активному бодрствованию (вплоть до так называемой «вегетативной бури»).
Выделяют две основные стадии сна - «медленный» и «быстрый» сон. Феномен «быстрого» сна открыли в 1953 г. американские ученые А. Азеринский и К. Клейтман. Медленноволновой сон человека периодически прерывается короткими периодами низкоамплитудной ЭЭГ (похожей на десинхронизацию ЭЭГ при пробуждении), сопровождающимися быстрыми движениями глазных яблок. Отсюда часто употребляемое название этой стадии - парадоксальная, или стадия быстрых движений глаз (БДГ-сон, или REM-стадия - от rapid eye movement). Люди, разбуженные во время парадоксальной стадии сна, в 80 % случаев сообщали о сновидениях. Мышцы тела расслаблены сильнее всего во время парадоксального сна, однако вегетативные показатели (дыхание, сердцебиение, кровяное давление и др.) во время парадоксального сна могут соответствовать активному бодрствованию (вплоть до так называемой «вегетативной бури»).

 График, показывающий возрастные изменения продолжительности и доли парадоксального сна у человека. Видно резкое сокращение длительности парадоксального сна (БДГ) - от 8 часов у новорожденных до 1 часа у пожилых людей. Изменения в продолжительности медленноволнового сна (МВ) не столь выражено; уменьшение от 8 часов до примерно 5 часов.
График, показывающий возрастные изменения продолжительности и доли парадоксального сна у человека. Видно резкое сокращение длительности парадоксального сна (БДГ) - от 8 часов у новорожденных до 1 часа у пожилых людей. Изменения в продолжительности медленноволнового сна (МВ) не столь выражено; уменьшение от 8 часов до примерно 5 часов.
 ЭЭГ человека при бодрствовании и сне Бодрствование Частота Амплитуда Синхронизация (регулярность колебаний) Медленноволновый сон Парадоксальный сон (БДГ) высокая низкая (нерегулярная активность, десинхронизация) высокая (синхронизированная активность) низкая (нерегулярная активность, десинхронизация)
ЭЭГ человека при бодрствовании и сне Бодрствование Частота Амплитуда Синхронизация (регулярность колебаний) Медленноволновый сон Парадоксальный сон (БДГ) высокая низкая (нерегулярная активность, десинхронизация) высокая (синхронизированная активность) низкая (нерегулярная активность, десинхронизация)
 В состоянии бодрствования с открытыми глазами преобладает бета-ритм, и этот же ритм наблюдается при парадоксальном сне. В процессе засыпания человек проходит 4 стадии, от спокойного бодрствования с закрытыми глазами (альфа-ритм) до собственно медленноволнового сна (дельта-ритм).
В состоянии бодрствования с открытыми глазами преобладает бета-ритм, и этот же ритм наблюдается при парадоксальном сне. В процессе засыпания человек проходит 4 стадии, от спокойного бодрствования с закрытыми глазами (альфа-ритм) до собственно медленноволнового сна (дельта-ритм).
 Основные ритмы ЭЭГ человека Частота Состояние 12 -25 Гц Бодрствование с открытыми глазами и БДГ-сон Альфа-ритм 8 -12 Гц Бодрствование с закрытыми глазами Тета-ритм 4 -8 Гц Засыпание 1 -4 Гц Медленноволновый сон Бета-ритм Дельта-ритм
Основные ритмы ЭЭГ человека Частота Состояние 12 -25 Гц Бодрствование с открытыми глазами и БДГ-сон Альфа-ритм 8 -12 Гц Бодрствование с закрытыми глазами Тета-ритм 4 -8 Гц Засыпание 1 -4 Гц Медленноволновый сон Бета-ритм Дельта-ритм
 ЭЭГ при различных уровнях бодрствования и сна. БДГ-сон напоминает по характеру ЭЭГ состояние бодрствования, однако электроды не регистрируют мышечной активности нигде, кроме глазных мышц.
ЭЭГ при различных уровнях бодрствования и сна. БДГ-сон напоминает по характеру ЭЭГ состояние бодрствования, однако электроды не регистрируют мышечной активности нигде, кроме глазных мышц.
 Препараты изолированного мозга (А) и изолированного переднего мозга (Б) с соответствующими записями ЭЭГ (так называемые «перерезки Бремера» - по Бремеру, 1937). При перерезке на границе головного мозга со спинным (препарат А) в ЭЭГ сохранялась картина бодрствования, а при перерезке на уровне среднего мозга (препарат Б) получали препарат спящего мозга. Бремер заключил, что в изолированном головном мозгу имеется центр пробуждения, расположенный в продолговатом и среднем мозгу.
Препараты изолированного мозга (А) и изолированного переднего мозга (Б) с соответствующими записями ЭЭГ (так называемые «перерезки Бремера» - по Бремеру, 1937). При перерезке на границе головного мозга со спинным (препарат А) в ЭЭГ сохранялась картина бодрствования, а при перерезке на уровне среднего мозга (препарат Б) получали препарат спящего мозга. Бремер заключил, что в изолированном головном мозгу имеется центр пробуждения, расположенный в продолговатом и среднем мозгу.
 Дж. Моруцци и Х. Мегун показали в 1949 г. , что стимуляция ретикулярной формации ствола мозга вызывает пробуждение. Сон при этом в то время рассматривался как следствие временной блокады активирующих восходящих влияний с одновременным «включением» таламокортикальных синхронизирующих процессов.
Дж. Моруцци и Х. Мегун показали в 1949 г. , что стимуляция ретикулярной формации ствола мозга вызывает пробуждение. Сон при этом в то время рассматривался как следствие временной блокады активирующих восходящих влияний с одновременным «включением» таламокортикальных синхронизирующих процессов.
 В настоящее время стало очевидно, что вместо единой «активирующей ретикулярной формации» (представление о которой сформировалось благодаря работам Бремера, Моруцци, Мегуна и других исследователей), существует большое количество центров, которые различаются по своим функциям, выделяемым нейромедиаторам и локализации в мозге. Большинство из них действительно находятся в ретикулярной формации ствола мозга или вблизи нее, но часть из них – в гипоталамусе и других структурах мозга.
В настоящее время стало очевидно, что вместо единой «активирующей ретикулярной формации» (представление о которой сформировалось благодаря работам Бремера, Моруцци, Мегуна и других исследователей), существует большое количество центров, которые различаются по своим функциям, выделяемым нейромедиаторам и локализации в мозге. Большинство из них действительно находятся в ретикулярной формации ствола мозга или вблизи нее, но часть из них – в гипоталамусе и других структурах мозга.
 Некоторые системы регуляции сна и бодрствования. Видно, как в цепях, выходящих из голубого пятна (медиатор - норадреналин) и ядер шва (медиатор - серотонин) аксоны расходятся к различным участкам спинного мозга, мозжечка и таламуса.
Некоторые системы регуляции сна и бодрствования. Видно, как в цепях, выходящих из голубого пятна (медиатор - норадреналин) и ядер шва (медиатор - серотонин) аксоны расходятся к различным участкам спинного мозга, мозжечка и таламуса.
 Выделение модулирующих медиаторов в кору больших полушарий во время бодрствования и сна Бодрствование Медленноволновый сон Парадоксальный сон Преобладающий ритм в ЭЭГ бета-ритм дельта-ритм бета-ритм Выделение норадреналина (из голубого пятна) максимально снижено отсутствует Выделение серотонина (из дорзальных ядер шва) максимально снижено отсутствует Выделение гистамина (из туберомамиллярного ядра заднего гипоталамуса) максимально снижено отсутствует Выделение ацетилхолина (из базальных ядер переднего мозга - базальное крупноклеточное ядро и др.) максимально отсутствует максимально
Выделение модулирующих медиаторов в кору больших полушарий во время бодрствования и сна Бодрствование Медленноволновый сон Парадоксальный сон Преобладающий ритм в ЭЭГ бета-ритм дельта-ритм бета-ритм Выделение норадреналина (из голубого пятна) максимально снижено отсутствует Выделение серотонина (из дорзальных ядер шва) максимально снижено отсутствует Выделение гистамина (из туберомамиллярного ядра заднего гипоталамуса) максимально снижено отсутствует Выделение ацетилхолина (из базальных ядер переднего мозга - базальное крупноклеточное ядро и др.) максимально отсутствует максимально
 В настоящее время очевидно, что бодрствование и сон – качественно различные состояния, каждое из которых имеет свои управляющие центры.
В настоящее время очевидно, что бодрствование и сон – качественно различные состояния, каждое из которых имеет свои управляющие центры.
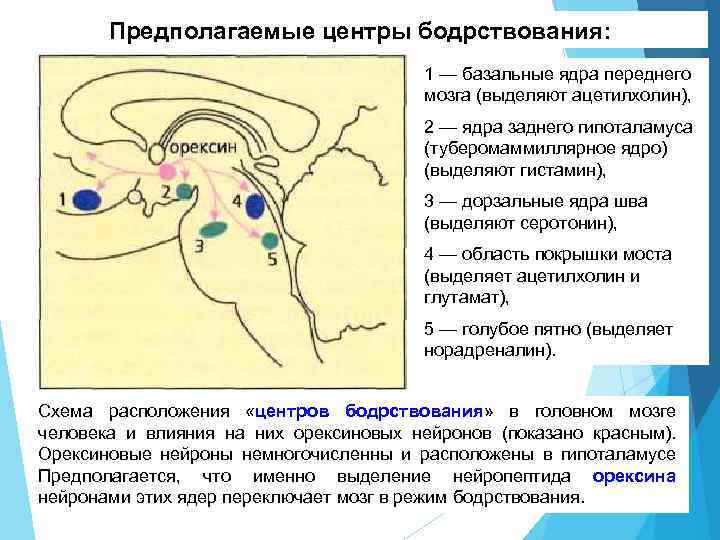 Предполагаемые центры бодрствования: 1 - базальные ядра переднего мозга (выделяют ацетилхолин), 2 - ядра заднего гипоталамуса (туберомаммиллярное ядро) (выделяют гистамин), 3 - дорзальные ядра шва (выделяют серотонин), 4 - область покрышки моста (выделяет ацетилхолин и глутамат), 5 - голубое пятно (выделяет норадреналин). Схема расположения «центров бодрствования» в головном мозге человека и влияния на них орексиновых нейронов (показано красным). Орексиновые нейроны немногочисленны и расположены в гипоталамусе Предполагается, что именно выделение нейропептида орексина нейронами этих ядер переключает мозг в режим бодрствования.
Предполагаемые центры бодрствования: 1 - базальные ядра переднего мозга (выделяют ацетилхолин), 2 - ядра заднего гипоталамуса (туберомаммиллярное ядро) (выделяют гистамин), 3 - дорзальные ядра шва (выделяют серотонин), 4 - область покрышки моста (выделяет ацетилхолин и глутамат), 5 - голубое пятно (выделяет норадреналин). Схема расположения «центров бодрствования» в головном мозге человека и влияния на них орексиновых нейронов (показано красным). Орексиновые нейроны немногочисленны и расположены в гипоталамусе Предполагается, что именно выделение нейропептида орексина нейронами этих ядер переключает мозг в режим бодрствования.
 Функция орексина как медиатора, ответственного за координацию других центров бодрствования, была открыта совсем недавно – около 2003 г. , а сам орексин был открыт лишь в 1998 г. При потере хотя бы части орексиновых нейронов возникает нарколепсия – дневная сонливость.
Функция орексина как медиатора, ответственного за координацию других центров бодрствования, была открыта совсем недавно – около 2003 г. , а сам орексин был открыт лишь в 1998 г. При потере хотя бы части орексиновых нейронов возникает нарколепсия – дневная сонливость.
 Схема центра расположения центра медленного сна. Вверху – бодрствование, когда центр сна заторможен и ретикулярная формация (РФ) активирует кору; внизу – центр сна возбужден, РФ заторможена и кора не активируется.
Схема центра расположения центра медленного сна. Вверху – бодрствование, когда центр сна заторможен и ретикулярная формация (РФ) активирует кору; внизу – центр сна возбужден, РФ заторможена и кора не активируется.
 Предполагаемые центры сна: Центр медленноволнового сна: в переднем гипоталамусе, (в вентролатеральном преоптическом ядре), медиатор - гаммааминомасляная кислота (ГАМК); активность нейронов незначительна в бодрствовании, но резко возрастает в период обычного сна и прекращается во время парадоксального. Центр парадоксального сна: ретикулярная формация покрышки моста (латеродорзальная / педункуло-понтийная области покрышки моста), медиаторы - ацетилхолин и глутамат.
Предполагаемые центры сна: Центр медленноволнового сна: в переднем гипоталамусе, (в вентролатеральном преоптическом ядре), медиатор - гаммааминомасляная кислота (ГАМК); активность нейронов незначительна в бодрствовании, но резко возрастает в период обычного сна и прекращается во время парадоксального. Центр парадоксального сна: ретикулярная формация покрышки моста (латеродорзальная / педункуло-понтийная области покрышки моста), медиаторы - ацетилхолин и глутамат.
 Список Литературы Дубынин В. А. Регуляторные системы организма человека. М. : Дрофа, 2003. Дудьев В. П. Психомоторика: cловарь-справочник, 2008 г. Мэгун Г. Бодрствующий мозг. М. : Мир, 1965. Росси Я. А. , Цанкетти А. Ретикулярная формация ствола мозга. М. : ИЛ, 1960. Шульговский В. В. Физиология центральной нервной системы. М. : Изд-во Моск. ун-та, 1987.
Список Литературы Дубынин В. А. Регуляторные системы организма человека. М. : Дрофа, 2003. Дудьев В. П. Психомоторика: cловарь-справочник, 2008 г. Мэгун Г. Бодрствующий мозг. М. : Мир, 1965. Росси Я. А. , Цанкетти А. Ретикулярная формация ствола мозга. М. : ИЛ, 1960. Шульговский В. В. Физиология центральной нервной системы. М. : Изд-во Моск. ун-та, 1987.
В основе регуляции смены активного и неактивного состояния животного организма лежат нервные и гуморальные процессы.
Для многих позвоночных животных определяющее значение в возникновении ритмов поведения имеет свет. Прежде всего это относится к птицам. У подавляющего большинства пернатых (исключение составляют ночные виды) с приближением вечерних сумерек развивается сонливость, дремота и сон. Утром на рассвете они просыпаются и включаются в активные действия. Такой же биоритм активности характерен и для млекопитающих животных с монофазной природой сна. У животных с полифазной природой сна связь с фотопериодом менее выражена или вообще отсутствует.
Усилиями физиологов, морфологов, биохимиков и гистохимиков доказано, что нервные клетки ядер шва обладают ритмической нейросекрецией. Снижение интенсивности афферентного потока со стороны экстеро-, интеро — и проприорецепторного аппарата организма животных приводит к падению неспецифической электрической активности ретикулярной формации ствола мозга, стимулирует функциональную активность ядер шва и позволяет неспецифическим ядрам таламуса синхронизировать электрическую активность коры.
Стимуляция ядер шва, в свою очередь, запускает процесс синтеза серотонина из аминокислоты триптофана. Образующийся в ядрах шва серотонин по аксонам нейронов распространяется к нейронам ретикулярной формации, таламуса, гипоталамуса, лимбической системы и блокирует их активирующее влияние на кору больших полушарий. Отсюда следует справедливое утверждение о том, что серотонин - это не просто медиатор центральной нервной системы, а медиатор сна . Однако его участие в механизме формирования сонного состояния специфично.
В специальных экспериментах было обнаружено, что искусственная блокада процессов синтеза серотонина устраняет у животных лишь фазу медленного сна и не влияет на фазу быстрого сна. Таким образом, серотонин выступает как посредник медленного сна .
Нейросекреторной активностью обладает еще одна область ствола мозга - так называемое синее пятно покрышки. Здесь вырабатывается норадреналин - медиатор пробуждения . Он выступает в качестве антагониста серотонина. Активность синего пятна, кроме того, приводит к торможению функциональной активности ядер шва. Другими словами, синее пятно покрышки и ядра шва находятся в реципрокных отношениях.
Интересную гипотезу регуляции ритма сна и бодрствования предложили американские исследователи А. Хобсон и Р. Мак-Карли (1977). В соответствии с их представлениями, биоритмы сна задаются спонтанной электрической активностью гигантских нейронов моста, которые имеют синаптические связи со многими структурами головного мозга. Ритмическая электрическая активность гигантоклеточного ядра моста, адресованная синему пятну, служит триггерным механизмом пробуждения. Активность гигантских нейронов моста, направленная на ядра шва, приводит к интеграции тормозных процессов и развитию сна. В данной схеме остаются неясными причины активизации тех или иных гигантских нейронов.
К настоящему времени оформились представления о существовании в мозгу двух систем, регулирующих сон и бодрствование.
Одна из них - восходящая активирующая ретикулярная система - расположена в верхних отделах ретикулярной формации ствола мозга и задних отделах гипоталамуса. При раздражении этой системы на электроэнцефалограмме появляется десинхронизация, уплощение и ускорение ритмов, что у спящих животных сопровождается пробуждением, а у бодрствующих повышением бдительности. Все периферические стимулы оказывают влияние на эту систему через коллатерали, которые отходят к ней от чувствительных путей, идущих к коре больших полушарий. Непосредственное электрическое раздражение корковых полей и некоторых других глубинных образований мозга может также вызвать пробуждение.
Но теперь уже очевидно, что раздражение всех отделов мозга, а также активность систем мозга, воспринимающих внешние и внутренние влияния, оказывают пробуждающее воздействие через восходящую активирующую систему. Опыты с перерезкой мозга и повреждением верхних отделов ретикулярной формации подтверждают это положение: животные погружаются в сонное состояние, из которого вывести их не удается.
Недавно появились сообщения, что при хорошем уходе у животных после операции через несколько недель появляются признаки бодрствования, увеличивающиеся со временем. Что это значит? Имеется ли еще одно звено активирующей системы, которое берет на себя осуществление этой функции, или в описанных опытах разрушается не вся восходящая система, сказать трудно. Возможным представляется существование активирующих аппаратов в лимбических структурах (миндалина, гиппокамп, таламус), функционально тесно связанных с аппаратами ретикулярной формации и гипоталамуса.
Более сложно построена вторая система - гипногенная, активность которой определяет длительность и глубину сна.
К настоящему времени уточнена роль ряда структур головного мозга в организации сна. Начнем с нижних отделов ствола. Моруцци описал синхронизирующий аппарат, при раздражении которого возникают электрофизиологические и поведенческие проявления сна. Роль этого образования сейчас хорошо выявлена: при отделении его (путем перезрезки) продолжительность сна у кошки уменьшается более чем в три раза. Животное бодрствует большую часть суток.
Разработан интересный способ анализа: в артерию вводят наркотическое вещество, выключающее временно функции определенных структур. Введение наркотика в сосуд, снабжающий нижний ствол кровью, приводит к тем же результатам, что и перерезка: удлиняется время бодрствования.
Этот аппарат тесно связан с каротидным синусом - образованием, расположенным в развилке наружной и внутренней сонных артерий, которое сигнализирует в мозг об уровне артериального давления и некоторых химических показателей. Раздражение каротидного синуса ведет к усилению деятельности синхронизирующего заднестволового аппарата, снятие раздражения - к обратному эффекту.
Роль барорецепторов этой зоны была подмечена уже давно, ведь не случайно артерии носят название «сонных». Известно, что в Индонезии на острове Бали знахари двухминутным массажем каротидного синуса вызывают сон. Совсем недавно французские нейрофизиологи описали в области нижнего ствола еще один синхронизирующий аппарат.
Другая гипногенная зона находится в области переднего гипоталамуса и перегородки. Раздражение этих структур электрическим током любой частоты приводит к синхронизации электроэнцефалографических ритмов и наступлению сна. Животное проделывает все ритуалы, характерные для его естественного сна (облизывание, мышечное расслабление, зевота). Разрушение этого аппарата приводит к длительному бодрствованию и резким нарушениям восстановительных процессов.
Еще одно важное звено в системе синхронизирующих аппаратов - таламическая синхронизирующая система. Раздражение низкочастотным электрическим током определенных ядер таламуса приводит к синхронизации потенциалов мозга и сну. Некоторые исследователи считают его главной гипногенной структурой, так как сон, наступающий при его раздражении, длителен и неотличим от нормального, а также вызывается легче, чем при раздражении других структур.
При низкочастотном раздражении сон можно вызвать, воздействуя на другие структуры мозга и даже периферические нервы. (Высокочастотное раздражение, как правило, приводит к пробуждению и десинхронизации.) Все это свидетельствует о распространенности синхронизирующих и десинхронизирующих аппаратов в нервной системе. Несомненно имеются сгущения, где они представлены более значительно. При разрушении этих скоплений и возникают эффекты противоположного характера - уменьшение или увеличение длительности сна.
Таким образом, имеются три главные гипногенные зоны, обеспечивающие возникновение и развитие сна. Мы знакомы с двумя видами сна, поэтому необходимо подчеркнуть, что эти структуры обеспечивают медленный сон. Как уже говорилось, за быстрый сон ответственны структуры средних отделов ствола мозга (ретикулярные ядра Варолиева моста). При их разрушении быстрый сон не наступает.
Гипногенная система по своей архитектуре сложна и включает многие аппараты мозга. Химически она, вероятно, неоднородна, так как в качестве медиаторов используются ацетилхолин, серотонин, гамааминомасляная кислота - ГАМК.
Что же представляет собой сон по своим физиологическим механизмам? Сразу же отпадает точка зрения, согласно которой сон - это отсутствие бодрствования, т. е. в основе его лежит выключение активирующих аппаратов. Очевидно, что имеются механизмы, организующие сон. Главное же заключается в том, что сон является активным организованным процессом, включающим различные по своей сущности и физиологическим механизмам состояния. Вот почему так сложно организована гипногенная система. Сон - сочетание активного состояния специализированных синхронизирующих аппаратов и снижения активности активирующей восходящей системы. Данные о состоянии отдельных нейронов во время сна хорошо подтверждают это положение. Следовательно, сами по себе отпадают представления о сне как охранительном, разлитом торможении. Только внешне это состояние можно охарактеризовать так. Однако приглядевшись, и в состоянии скелетно-мышечной системы можно видеть активность. Интенсивная психическая деятельность во время сна также говорит об активности мозга в этом состоянии.
Итак, существуют две системы, регулирующие сон и бодрствование. У систем имеются подсистемы, включающие различные формы сна в определенной последовательности. Все наводит на мысль о существовании в мозгу координирующего аппарата, который в определенное время регулирует включение отдельных систем в целом, а затем и их подсистем. В этом нас убеждают наблюдения над больными людьми, когда все подсистемы работают, но резко нарушается закономерная последовательность их включения. Координирующий аппарат не находится в одном каком-либо отделе мозга. Речь идет о сложном комплексе с преимущественным расположением в передних отделах больших полушарий мозга, лимбических аппаратах, гипоталамусе. Дальнейшие исследования позволят более четко и обоснованно представлять подобную точку зрения.
Освещая современные представления о регуляции сна и бодрствования, нельзя не вернуться к гуморальным факторам в происхождении сна. Поиски гипнотоксинов - веществ, накопление которых вызывает сон, ведутся очень давно. Имеется целый ряд исследований, результаты которых трудно объяснить без участия какого-то гуморального агента. В исследовании немецкого физиолога Кроля показано, что экстракт вещества головного мозга спящего животного при внутривенном введении его вызывает сон у подопытного животного. Выше уже были описаны опыты (Монье, Корнмюллера): у подопытного животного наступал сон, если в его организм поступала кровь из мозга другого животного, погруженного в сон в результате раздражения зрительного бугра.
Известным физиологом А.В. Тонких была показана роль гормонов, главным образом гипофиза, в возникновении сна. В Лаборатории по изучению нервных и гуморальных регуляций имени Н.И. Гращенкова также проведены специальные исследования содержания активных биологических веществ в крови и моче у больных с повышенной сонливостью. Было установлено, что содержание в крови и в моче адреналина (гормонов коркового слоя надпочечника) снижено, а ацетилхолина, гистамина и продуктов обмена серотонина повышено.
Эти данные несомненно интересны, однако возникает вопрос, насколько содержание биологически активных веществ на периферии отражает их истинные соотношения в головном мозге. На примере сиамских близнецов опровергается гуморальная теория возникновения сна. Следует иметь в виду, что в циркулирующих жидкостях содержание активных биологических веществ одинаково, а в мозгу различно. Возможно, именно поэтому одна голова спала, а другая в это время бодрствовала. Вот почему в последние годы стали уделять особое внимание химическим передатчикам нервных импульсов, богато представленным в мозге.
В настоящее время очевидно присутствие в мозге медиаторов - веществ, выделяющихся в синапсах на границе двух нейронов и обеспечивающих распространение нервного импульса. Холинэргические синапсы в качестве медиатора выделяют ацетилхолин, адренэргические - норадреналин, серотонинэргические - серотонин. Есть синапсы с гамма-аминомасляной кислотой и большое число синапсов с еще не идентифицированным химическим передатчиком. Все химически неоднородные нейроны не разбросаны в мозгу хаотично, а составляют определенные системы, объединяемые по принципу представленности в них того или иного медиатора. Норадреналин и серотонин обнаруживаются главным образом в глубинных и стволовых структурах мозга, в то время как ацетилхолин распределен более равномерно.
Различным химическим системам придавалось и определенное функциональное значение. На опытах доказано, что активирующая восходящая ретикулярная система, поддерживающая необходимый уровень бодрствования, по своей химической характеристике адренэргическая, что введение адреналина усиливает настороженность животного, а во сне содержание его в мозгу снижается. Многие фармакологические средства, препятствующие сну, близки по составу к адреналину либо, вмешиваясь в химию мозга, способствуют накоплению этих веществ. Правда, установлено, что нередко одна функциональная система является гетерохимической, т. е. в нее входят нейроны, медиаторы, которые различны по химическому составу.
В последнее время формируется представление, согласно которому основные гипногенные вещества - ацетилхолин, серотонин и ГАМК. Физиолог из Южной Америки Эрнандец-Пеон обнаружил с помощью специальных опытов, что наложение кристаллика ацетилхолина на структуры ствола мозга, гипоталамуса, медиальных отделов височной доли вызывает электроэнцефалографические и поведенческие признаки сна. Косвенным доказательством является и то, что в отделах мозга, где расположены гипногенные аппараты, основной медиатор - ацетилхолин.
Накапливаются также факты, которые говорят о роли серотонина. Разрушения ядер шва, расположенных в стволе мозга и наиболее богатых серотонином, приводят к бессоннице, степень которой обратно пропорциональна числу сохраненных ядер. Проведены многочисленные опыты с введением в организм аминокислоты - предшественника серотонина - триптофана и антагонистов серотонина - метисергида, дезерила, разрушающих его и оказывающих противоположное действие на сон в целом и на его отдельные фазы. По этому вопросу возникла дискуссия: одни ученые отстаивают мнение, согласно которому серотонин способствует возникновению быстрого сна, другие - медленного. По-видимому, более обоснованна вторая точка зрения. Удалось показать, что адренэргические аппараты участвуют не только в механизмах бодрствования, но и быстрого сна. Эти исследования имеют огромное практическое значение, так как являются базой для создания современной дифференцированной фармакологии сна и бодрствования.
Перспективы этой проблемы заключаются скорее всего не в поисках каких-то особых гипногенных веществ, а в выяснении истинной роли уже известных химических активных агентов и в идентификации еще не опознанных медиаторов головного мозга.
Интерес к гуморальным исследованиям особенно обострился в связи с открытием быстрого сна. Было установлено, что лишение людей и животных быстрого сна приводит к увеличению этой фазы сна в последующие ночи. Создалось впечатление, что в фазе быстрого сна разрушается какое-то гипотетическое вещество, накапливающееся во время бодрствования. Следовательно, при лишении быстрого сна этот фактор продолжает накапливаться и в последующие ночи вызывает избыточную длительность этой фазы. Однако против подобной гипотезы говорит ежедневное наблюдение, связанное с большей продолжительностью быстрого сна во второй половине ночи, когда накопившееся вещество должно было бы уже разрушиться. Тем не менее такими общими, хотя и логическими соображениями противостоять указанной гипотезе трудно. Нужны факты. Пока же создается впечатление, что отдельные фазы сна имеют собственную химию.
А. Сон - это особая активность мозга, при которой у человека выключаются сознание и механизмы поддержания естественной позы, снижена чувствительность анализаторов. Рекомендуе мая продолжительность сна взрослого человека 7-8 часов в сутки.
Для оценки глубины сна обычно используют электроэнцефалограмму (ЭЭГ). По особенностям ЭЭГ, исходя из общепринятых стандартных критериев, выделяют четыре или пять его стадий (рис. 13.8).
Перед пробуждением спящий человек обычно проходит через особую фазу сна, характеризующуюся десинхронизацией ЭЭГ и эпизодами быстрых движений глаз (БДГ). Их можно наблюдать со стороны через сомкнутые веки спящего или записывать методами электроокулографии. БДГ настолько характерны для данной фазы, что ее называют сном с БДГ - быстрым сном, другие фазы называют медленным (синхронизированным) сном. Остальная мускулатура в фазе быстрого сна, как и во время медленного сна, практически атонична, если не считать иногда возникающих судорожных сокращений мышц лица или пальцев.
Порог пробуждения в БДГ-фазе сна примерно такой же, как и во время глубокого сна, однако ЭЭГ сходна с записываемой при бодрствовании или переходе ко сну, поэтому БДГ-сон получил также название парадоксального, или десинхронизированного.
На протяжении ночи последовательность стадий сна повторяется в среднем три - пять раз. Как правило, максимальная его
глубина при каждом таком цикле убывает к утру. В норме БДГ-сон повторяется примерно через каждые 1,5 ч и длится в среднем 20 мин, причем с каждым разом все больше.
Б. Сновидения - возникающие во сне образные представления, воспринимаемые как реальная действительность. Сновидения, очевидно, возникают главным образом во время сна с БДГ.
Факторы, побуждающие сновидения. 1. Предшествующая сну деятельность (дети продолжают «играть» во сне, исследователь ставит эксперименты и т. д.). Например, известному физиологу О. Леви приснилась модель опыта, с помощью которого он открыл медиаторный механизм передачи влияний с симпатического и парасимпатического нервов на сердце. Д. И. Менделееву сновидение помогло создать свою знаменитую таблицу. 2. Раздражители, действующие на организм во время сна. Так, если приложить горячую грелку к ногам спящего человека, может присниться сон, что он идет по раскаленному песку. 3. Избыточная импульсация от переполненных или больных внутренних органов может вызывать кошмарные сновидения.
В. Значение сна.
Сон обеспечивает отдых организма. В экспериментах М. М. Манассеиной (1892) было показано, что лишенные сна взрослые собаки погибали на 12-21-й день. Лишение сна щенков приводило их к гибели через 4-6 дней. Депривация сна человека в течение 116 часов сопровождалась нарушением поведения, повышением раздражительности, психическими расстройствами. Более значительно меняется поведение человека при лишении его медленного сна, в результате чего возникает повышенная возбудимость.
Сон играет важную роль в процессах метаболизма. Полагают, что медленный сон способствует восстановлению внутренних органов, поскольку через гипоталамус либерины воздействуют на гипофиз, способствуя освобождению гормона роста (ГР), который участвует- в биосинтезе белков в периферических тканях. Напротив, парадоксальный сон восстанавливает пластические свойства нейронов головного мозга, усиливает процессы в клетках нейроглии, которые обеспечивают нейроны питательными веществами и кислородом.
3. Сон способствует переработке и запоминанию информа ции. Информация, предъявляемая во время сна, не запомина ется, если только на ЭЭГ во время или после этого не появляется а-ритм (т. е. если человек не просыпается). Из всех проявлений активности мозга во время сна запоминается лишь последнее сно видение. С другой стороны, сон облегчает закрепление изучен-
ново материала. Если какая-то информация заучивается непосредственно перед засыпанием, то спустя 8 ч она вспоминается лучше (утро вечера мудренее).
4. Биологическое значение сна связано с приспособлением к изменению освещенности (день-ночь). Организм способен заранее приспособиться к ожидаемому воздействию со стороны внешнего мира, активность всех систем снижается в определенные часы согласно режиму труда и отдыха. К моменту пробуждения и в начале бодрствования активность органов и систем возрастает и соответствует уровню поведенческих реакций.
Г. Механизмы бодрствования и сна.
Переход от бодрствования ко сну предполагает два возможных пути. Прежде всего, не исключено, что механизмы, поддерживающие бодрствующее состояние, постепенно «утомляются». В соответствии с такой точкой зрения, сон - это пассивное явление, следствие снижения уровня бодрствования. Однако не исключено и активное торможение обеспечивающих бодрствование механизмов. Важную роль в регуляции цикла сон - бодрствование играет ретикулярная формация ствола мозга, где находится множество диффузно расположенных нейронов, аксоны которых идут почти ко всем областям головного мозга, -за исключением неокортекса. Ее роль в цикле сон - бодрствование была исследована в конце 1940-х гг. Г. Моруцци и Н. Мэгуном. Они обнаружили, что высокочастотное электрическое раздражение этой структуры у спящих кошек приводит к их мгновенному пробуждению. И напротив, повреждения ретикулярной формации вызывают постоянный сон, напоминающий кому; перерезка же только сенсорных трактов, проходящих через ствол мозга, такого эффекта не дает.
Серотонинергические нейроны также играют весьма важную роль в регуляции бодрствования и сна. В верхних отделах ствола мозга есть две области - ядра шва и голубое пятно, у нейронов которых такие же обширные проекции, как и у нейронов ретикулярной формации, т. е. достигающие многих областей ЦНС. Медиатором в клетках ядер шва служит серотонин (5-гидрокситрипта-мин, 5-НТ), а голубого пятна - норадреналин. Разрушение ядер шва у кошки приводит к полной бессоннице в течение нескольких дней; за несколько следующих недель сон нормализуется. Частичная бессонница может быть также вызвана подавлением синтеза 5-НТ я-хлорфенилаланином. Ее можно устранить введением 5-гид-рокситриптофана, предшественника серотонина (последний не проникает через гематоэнцефалический барьер). Двустороннее разру-
шение голубого пятна приводит к полному исчезновению БДГ-фаз, не влияя на медленноволновой сон. Истощение запасов серотонина и норадреналина под влиянием резерпина вызывает, как и следовало ожидать, бессонницу.
Были сделаны попытки обнаружить особые вещества либо после длительного лишения сна, либо у спящего человека. Первый из этих подходов основан на предположении о том, что фактор(ы) сна во время бодрствования накапливаются до вызывающего сон уровня, а второй - на гипотезе, согласно которой они образуются или выделяются во сне.
Оба подхода дали определенные результаты. Так, при проверке первой гипотезы из мочи и спинномозговой жидкости человека и животных был выделен небольшой глюкопептид - фактор 5, вызывающий медленноволновой сон при введении другим животным.
Однако сросшиеся девочки-близнецы могли спать порознь, что свидетельствует о второстепенной роли гуморальных факторов и решающей роли нервной системы в развитии сна.
Переход от бодрствования ко сну предполагает два возможных пути. Прежде всего не исключено, что механизмы, поддерживающие состояние бодрствования, постепенно «утомляются». В соответствии с такой точкой зрения сон - это пассивное явление, следствие снижения уровня бодрствования. Однако не исключено и активное торможение механизмов, обеспечивающих бодрствование. И.П. Павлов выделял два механизма развития сна, которые, по существу, подтверждают правомерность позиций сторонников как пассивной, так и активной теории сна. С одной стороны, сон возникает как явления охранительного торможения в результате сильного и длительного раздражения какого-либо отдельного участка коры больших полушарий. С другой стороны, сон возникает как результат внутреннего торможения, т.е. активного процесса формирования отрицательного условного рефлекса. Важную роль в регуляции цикла сон – бодрствование играет ретикулярная формация ствола мозга, где находится множество диффузно расположенных нейронов, аксоны которых идут почти ко всем областям головного мозга, за исключением неокортекса. Роль РФ в цикле сон – бодрствование была исследована в конце 1940-х годов учеными Г. Моруцци и Н. Мэгуном, обнаружившими, что высокочастотное электрическое раздражение этой структуры у спящих кошек приводит к их мгновенному пробуждению. И напротив, повреждения ретикулярной формации вызывают постоянный сон, напоминающий кому; перерезка же только сенсорных трактов, проходящих через ствол мозга, такого эффекта не дает. Наиболее ранними теориями сна были гуморальные. Фактор сна, лишенный видовой специфичности, был выделен из ликвора коз, подвергнувшихся депривации сна. Согласно сосудистой (циркуляторной или гемодинамической) теории сна, наступление сна связано со снижением кровотока в мозге или с его усилением. Современные исследования показали, что в течение сна действительно происходит колебание кровенаполнения мозга. Р. Лежандр и X. Пьерон (1910) считали, что сон возникает в результате накопления токсических продуктов обмена вследствие утомления (гипотоксины). Собакам долгое время не давали спать, а затем забивали, экстрагировали вещества из мозга и вводили другим собакам. У последних развивались признаки крайнего утомления и возникал глубокий сон. То же наблюдалось при «переносе» сыворотки крови или спинно-мозговой жидкости.
В 1965 г. Ж. Монье использовал препарат с перекрестным кровообращением у двух кроликов, т.е. кровь от мозга одного кролика попадала в туловище другого. Если у одного из кроликов раздражали участки мозга, вызывающие сон, то второй кролик тоже засыпал. Во время сна в мозгу обнаружено избыточное накопление ряда биологически активных веществ – ацетилхолина, гаммааминомасляной кислоты, серотонина.
В середине XIX в. большее распространение получили нервные теории сна. Один путь исследований – это изучение так называемого центра сна, о существовании которого в ядрах гипоталамуса у животных утверждал швейцарский физиолог В. Гесс (1933). У больных с пораженным гипоталамусом также отмечалась повышенная сонливость: у раненого солдата осколок снаряда находился на уровне гипоталамуса, а попытка извлечь осколок пинцетом вызывала мгновенный глубокий сон.
Выделяют так называемую информационную теорию сна (Н. Вингер). Согласно этой теории, в течение дня мозг накапливает огромную информацию, усвоение которой затруднено, а часть ее не имеет отношения к долговременным задачам. Если кратковременная память заполняется днем, то ночью часть содержащейся в ней информации переписывается в долговременную память. Особенности процессов переработки информации требуют отключения от сигналов внешнего мира.
В верхних отделах ствола мозга есть две области – ядра шва и голубое пятно, у нейронов которых такие же обширные проекции, как и у нейронов ретикулярной формации, т.е. достигающие многих областей ЦНС. Ядра шва захватывают срединную часть продолговатого мозга, моста и среднего мозга. Разрушение их устраняет синхронизацию ЭЭГ и медленный сон. С помощью специальной методики флуоресценции гистохимики показали, что нейроны ядер шва синтезируют серотонин и направляют его через свои аксоны к ретикулярной формации, гипоталамусу, лимбической системе. Серотонин – тормозной медиатор моноаминергической системы мозга. Блокада синтеза серотонина устраняет у кошки медленный сон, у которой сохраняется лишь парадоксальный сон.
В среднем мозге (покрышка) обнаружено скопление нейронов, синтезирующих норадреналин (голубое пятно). Стимуляция голубого пятна вызывает торможение нейронной активности во многих структурах мозга при росте двигательного возбуждения животного и ЭЭГ-десинхронизации. Полагают, что активирующее влияние голубого пятна осуществляется через механизм торможения тормозных интернейронов. Ядра шва и голубое пятно действуют как антагонисты. Медиатором в клетках ядер шва служит серотонин (5-гидрокситриптамин, 5-НТ), а голубого пятна – норадреналин. Разрушение ядер шва у кошки приводит к полной бессоннице в течение нескольких дней; но за несколько последующих недель сон нормализуется. Частичная бессонница может быть также вызвана подавлением синтеза 5-НТ п -хлорфенилаланином. Ее можно устранить введением 5-гидрокситриптофана, предшественника серотонина (последний не проникает через гематоэн-цефалический барьер). Двустороннее разрушение голубого пятна приводит к полному исчезновению БДГ-фаз, не влияя на медленноволновый сон. Истощение запасов серотонина и норадреналина под влиянием резерпина вызывает, как и следовало ожидать, бессонницу. Однако оказалось, что нейроны ядер шва наиболее активны и выделяют максимум серотонина не во время сна, а при бодрствовании. Кроме того, возникновение БДГ, по-видимому, обусловлено активностью нейронов не столько голубого пятна, сколько более диффузного подголубого ядра. Судя по результатам недавних экспериментов, серотонин служит и медиатором в процессе пробуждения, и «гормоном сна» в бодрствующем состоянии, стимулируя синтез или высвобождение «веществ сна» (факторов сна), которые в свою очередь вызывают сон. Структуры таламуса выполняют функцию «пейсмекера» для вызова ритмических потенциалов веретен во сне и a-ритма в бодрствовании. Таламокортикальный механизм можно рассматривать как механизм внутреннего торможения, способного изменять активность мозга частично или глобально таким образом, что сенсорные, моторные и высшие функции мозга подавляются.
Рис. 9.4. Реципрокные отношения мультинейронной активности ретикулярной формации и преоптической области гипоталамуса в различных фазах цикла бодрствование–сон; калибровка: 40 мкВ, 50 мс (по Т.Н. Ониани, 1983, с изменениями)
Структуры, ответственные за медленный сон, находятся в каудальной части мозгового ствола, главным образом – в продолговатом мозге. Наличие сходных гипногенных структур было установлено также и в задней части моста. Двигательные и ЭЭГ-проявления фазы парадоксального сна связаны с активацией структур в области моста. Эта фаза сна сокращается при эмоциональном стрессе, при этом удлиняется период засыпания.
Рядом с голубым пятном имеется группа гигантских ретикулярных нейронов, которые направляют свои аксоны вверх и вниз к различным структурам мозга. В бодрствовании и медленном сне эти нейроны малоактивны, но их активность весьма высока во время парадоксального сна.
Были сделаны попытки обнаружить особые вещества либо после длительного лишения сна, либо у спящего человека. Первый из этих подходов основан на предположении о том, что фактор(ы) сна во время бодрствования накапливаются до вызывающего сон уровня, а второй – на гипотезе, согласно которой они образуются или выделяются во сне.
Оба подхода дали определенные результаты. Так, при проверке первой гипотезы из мочи и спинномозговой жидкости человека и животных был выделен небольшой глюкопептид – фактор S, вызывающий медленноволновый сон при введении другим животным. Существует, по-видимому, и фактор сна с БДГ. Второй подход привел к открытию индуцирующего глубокий сон нонапептида (в настоящее время он уже синтезирован), так называемого пептида D-сна (DSIP , delta-sleep inducing peptide). Однако пока неизвестно, играют ли эти и многие другие «вещества сна», обнаруженные при проверке обеих гипотез, какую-либо роль в его физиологической регуляции. Более того, выделенные пептиды часто вызывают сон лишь у животных определенного вида; кроме того, он возникает и под действием других веществ.
Однако сросшиеся девочки-близнецы могли спать порознь, что свидетельствует о второстепенной роли гуморальных факторов и решающей роли в развитии сна нервной системы.
Развивается представление о том, что цикл бодрствование – сон обеспечивается системой двух центров. К. Экономо на основе клинических наблюдений больных с повреждениями различных участков гипоталамуса предположил, что центр бодрствования локализован в заднем, а центр сна – в его передних отделах. С. Рэнсон, производя локальные повреждения различных участков гипоталамуса, подтвердил это мнение. В настоящее время считают, что гипоталамус является критической зоной для регулирования цикла бодрствование – сон. Это мнение подтверждается и тем, что как высокочастотное, так и низкочастотное электрическое раздражение преоптической области гипоталамуса вызывает синхронизацию электроэнцефалограммы и поведенческий сон. Противоположный эффект, а именно поведенческое и электроэнцефалографическое пробуждение Т.Н. Ониани наблюдал при раздражении заднего гипоталамуса. Это позволяет предположить наличие реципрокного взаимоотношения между передней и задней областями гипоталамуса и его значение для регуляции чередования различных фаз цикла бодрствование – сон. По данным Т.Н. Ониани, в цикле бодрствование – сон мультинейронная активность ретикулярной формации (активирующей системы) и преоптической области гипоталамуса (тормозной системы) меняется реципрокно (рис. 9.4).
Гипноз
Гипноз – состояние человека, вызванное искусственно с помощью внушения, когда не полностью выключено сознание и снижены реакции на все раздражители, кроме слов гипнотизирующего, к которым человек становится чрезвычайно восприимчивым. В ответ на многократные или монотонные стимулы непременно развивается внутреннее торможение. Если такая стимуляция продолжается, то наступает сон. Переходный период между бодрствованием и сном назван гипнотическим состоянием. И.П.Павлов считал, что гипнотическое состояние является частичным сном. Он разделил гипнотическое состояние на три фазы.
Первая фаза называется уравнительной, в это время сильные и слабые стимулы вызывают одинаковые условные ответы.
Парадоксальная фаза характеризуется более глубоким сном, слабые раздражители вызывают более интенсивный ответ, чем сильные.
Ультрапарадоксальная фаза означает еще более глубокий сон, когда ответ вызывают только слабые стимулы, а сильные приводят к еще большему распространению торможения. За этими тремя фазами следует глубокий сон.
Однако оказалось, что в гипнозе волны электроэнцефалограммы (ЭЭГ) сходны с ЭЭГ бодрствования, а не сна. Выяснилось, что одни люди более подвержены гипнозу, чем другие, а некоторые вовсе не поддаются ему (что говорит о различном соотношении процессов возбуждения и торможения у разных людей). В последние годы получили распространение теории, согласно которым гипнотическое состояние нельзя приравнивать ни ко сну, ни к бодрствованию.
Обычно выделяют четыре стадии гипноза.
1. Стадия гипноидности, сопровождающаяся психическим и мышечным расслаблением, миганием и закрыванием глаз.
2. Стадия легкого транса, для которой характерна каталепсия конечностей (конечности могут длительное время находиться в необычном положении).
3. Стадия среднего транса, когда возникает амнезия, изменения личности, возможны простые гипнотические внушения.
4. Стадия глубокого транса, которая характеризуется полным сомнамбулизмом и фантастическими внушениями.
При гипнозе изменяется состояние коры больших полушарий, которое меняет соотношение между первой и второй сигнальными системами. Снижается контроль второй сигнальной системы над первой и облегчается воспроизведение пережитых состояний и зрительных образов. Возбуждение коры при восприятии слов гипнотизера оказывается изолированным от влияния остального мозга, воздействия на него оказываются необычайно сильными с длительным последействием, что обусловливает и длительный лечебный эффект гипнотического внушения (гипнотерапии).
Во время гипноза высшая нервная деятельность существенно меняется. Это касается сенсорной сферы, когда можно внушить отсутствие слуха, зрения, осязания и даже боли, можно получить извращенные реакции (иллюзии), например на вкусовые или обонятельные раздражения, вызвать галлюцинации – восприятие несуществующих предметов и явлений (положительные) или выключить из восприятия определенные предметы (отрицательные галлюцинации). Принципиальный интерес представляют исследования с внушением возраста, когда у человека восстанавливаются особенности памяти, речи, почерка, характерные для его детства, меняется фонетическая выразительность речи, темп и содержание мышления.
Гипнотику можно внушить различные эмоциональные состояния, воздействовать на интеллектуальные, моральные и этические чувства, на сферу его внимания, но нельзя изменить личность человека вопреки его социальным и индивидуальным убеждениям.
Механизмы гипноза остаются неизвестными, а завершенной теории гипноза еще не разработано, хотя ряд заболеваний успешно излечивается под гипнозом.


